An ecomorphic
theatre as a case study for embodied design*
Alasdair Turner,
MA, MSc
Bartlett School of Graduate Studies, University
College London, London WC1E 6BT
e-mail:
a.turner@ucl.ac.uk
Abstract
In this paper we examine the ecomorphic design of a
‘theatre’. The theatre is defined
through the relationships of the players and audience. The players occupy a fixed area, and the
audience may sit around them. Each
member of the audience must be able to access their seating position, and, once
seated, each is given score based on how well they may view the players. After the conditions have been set down, we
attempt to evolve the theatre to make a good fit for the audience and players. The idea is to create an embodied system
which captures the interplay between visibility and accessibility. We compare several different generative
techniques to create the theatre. In
the first place, we attempt a continuous process whereby each audience member
themselves may raise or lower a column in order to view the play. Then we allow the theatre itself to evolve,
based on live cell growth from its edges, so it becomes a cellular automata
system guided by the visibility and accessibility of its growth. We also apply basic forms of
diffusion-limited aggregation, genetic algorithms and genetic programs. We find that the cellular growth model
provides the best solutions in terms of number of accessible seating positions
and the best visibility for each audience member, whilst also creating the most
interesting design outcomes. We suggest
that this is because the cellular growth model naturally suits ecomorphic
principles of design through continuous adjustment of the environment around
its occupation to meet a natural interaction between audience and players.
1. Introduction
Ecomorphic design was introduced by the author in
2002 [1], in order to tie together a number of concepts centered around
embodiment, situatedness and generative design. It might best be expressed as the process of structural coupling
as described by Maturana and Varela [2].
Originally, Maturana and Varela presented the idea of structural
coupling in the domain of cell biology and within the domain of their theory of
autopoiesis. They considered cells as
autopoietic, that is, cell maintaining within the environment. Varela et al went on to extend these ideas
into the domain of cognitive science, and specifically embodiment [3]. To Varela, following Heidegger, the mind is
embodied, and its actions and reactions may take place only in the context of
its environment. To Varela et al,
embodiment is the middle path between the Cartesian dualist mind body split,
and the Liebnizian view of a single monadology. Varela et al at once see the mind-body unit as separate from its
environment, but simultaneously, as only possibly existing within that
environment (that is, the being is meaningful only in relation to
environment). Returning to Maturana and
Varela’s original conception, structural coupling occurs when the environment
and the cell mould their physical form to each other. Varela et al’s being would also be structurally coupled through
physical interaction with it (we might compare Thompson’s model of bone growth,
where the being creates bone in response to the physical demands of the
environment [4], or patterns of stigmergy, where the environment is shaped
through its occupation by beings [5]).
Ecomorphism builds on this theoretical basis
primarily through Luhmann [6]. Rather
than considering directly the coupling of environment and a single being,
ecomorphism considers the coupling of a social process and the
environment. Luhmann suggests that the
ongoing process of the being and the ongoing process of the social phenomena
are both autopoietic in nature, and can be considered simply in terms of
whether they are psychic (concerning the individual) or social (a collection of
individuals). Thus ecomorphism is still
concerned with the coupling of the environment to a being, albeit a distributed
social process being.
In this paper, we examine the case of an ecomorphic
theatre. The social process to be
engaged in is a performance. We shall
use a relatively standard interpretation of this process, with the audience
engaged in viewing the actors, rather than some further interaction between
them. The process will concern the
growth of the environment to support this activity. For ease of presentation, the seating area will be defined
outside the actors’ stage space, and it is this seating area that will be
modified. Strictly speaking, the
ecomorphic environment should have no predefined distinction between stage and
auditorium such as this: the actors and audicence should be free to arrange
themselves within the environment, and the environment should evolve around
this activity. If this means the actor
stands atop a mountain and the audience look up to her or him, then this is a
valid outcome. However, we should be
clear that the action of looking and
the ability to move to a place so as
to view are fundamental to the ecomorphic process and cannot be excluded. In considering these factors, we set up an
interplay between accessibility of a space and visibility to and from that
space. This reflects the theory of
space syntax [7], which examines how these variables may interact to structure
space and the social process within it.
However, whereas space syntax is primarily analytic, in this paper, we
will try to examine this interplay through generation of a space.
2. Background
The techniques used to generate morphology in this paper
have generally been used extensively within the field of generative
architecture [e.g.,8,9]. In the case of
each algorithm, the same evaluation function will be used, described in section
2.2, either for the members of the audience as a whole, or for individuals
seated within it, depending on the algorithm.
2.1 Generative algorithms overview
We first implement an individual based algorithm:
each audience member chooses a location, and may build on the location to
obtain a better view. This then leads
on to a less individualistic interpretation, with the audience members being
collectively rewarded for achieving the best view for everyone. However, both these methods are restricted
to a set number of audience members: we define a 20 x 20 grid of boxes, with a
5 x 5 area for the stage, leaving 375 places to sit. If we want to increase the number of seats, we must allow the
columns to grow outwards, into the space.
This rule thus becomes a form of 3D cellular automata system [10]. The rules for when to grow an adjoined
structure depend on whether or not the population get a better view, or through
adding more audience members, thus increasing the population numbers with any
view at all. We then moved on to test
other forms of well know generative algorithm: diffusion limited aggregation
[11], a straight-forward genetic algorithm using a direct translation from
genotype to phenotype [12], and then a genetic program, allowing different
types of cell to grow outwards from a seed [13].

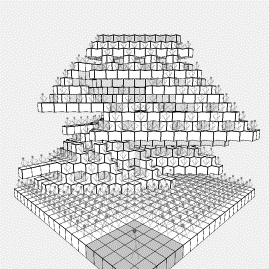
Figure
1: a typical system
evolved using the cellular growth model
2.2 The reward function
All the algorithms are evaluated using the same
reward function, which combines accessibility and visibility. A typical proposal to be evaluated is shown
in figure 1. To give an idea of the
intended scale, each block is 75 x 75 x 75cm, with a growth space of 20 x 20 x
20 blocks (15m in all dimensions). We
first evaluate which locations are accessible. These are discovered by starting with the
location of the actor shown on the stage.
A location is accessible if it is adjacent to another accessible
location, and if it is either at the same height as the accessible location,
one block higher, or one block lower (either step up 75cm or down 75cm). In addition, there must be a head-clearance
of two blocks (1.5m) to allow transit between locations. Once all the accessible locations have been
discovered using a breadth-first search, we then assess the visibility from each
of these locations. An audience member
may sit at the accessible location if she or he has an unobstructed view of the
upper torso and head of the actor. The
visibility is assessed by pixelating the path from the viewer to the actor at
the scale of the blocks. The audience
member is rewarded according to the number of other audience members obscuring
the view of the actor. There is a
maximum reward of 20, with 1 deducted for each member of the audience in the
(pixelated) line of vision to the upper torso and head of the actor. Before any evolution of the system, this
works out at an average of about 6.6 other audience members obscuring the view.
3. Experiments
As discussed above, we start with an individualistic
algorithm. Each audience member finds a
location, and the block is raised according to whether she or he personally
benefits from the change. At the
beginning of this process, the audience are evenly distributed over the grid of
blocks, as shown in figure 2(a).

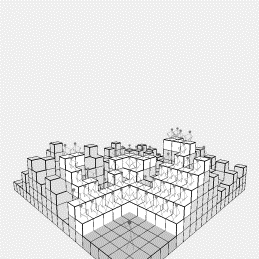
Figure
2: (a) Early growth
of individualistic agents (b) Later growth: a few agents block other locations
from seeing
However, as the system progresses, agents near the
front block other agents from viewing, and as the system grows, raise up the
blocks so that other locations within the system are no longer accessible
(starting from the stage as discussing in section 2.2). Figure 2(b) shows the converged end product
of a run. There are just 52 audience
members remaining, from the 375 that started, although each has a completely
unobscured view of the actor. In order
to overcome this problem, we moved to a second form of algorithm, where columns
are raised, but only if the overall fitness of the system is increased by doing
so. As might be expected, the columns
start rising from the back, so they do not block those at the front, as shown
in figure 3(a). As the system run
progresses, the seats nearer the back rise higher to compensate for the angle
of view, as shown in figure 3(b). The
end result is that all 375 audience members can view, mostly unobscured by
others.

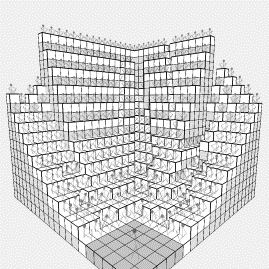
Figure
3: (a) Early growth
of the social optimum system starts from the back (b) a later result, with
seating rising faster to the rear
This system however lacks interest, and can only ever
reach an optimum based on 375 audience members. In order to increase the viewing audience, we allowed any
accessible space to grow not only upwards, but outwards (in any direction),
with the results shown in figure 4. The
growth starts as before, but after a while the higher cells start to grow
towards the stage, allowing more audience members to view the stage. The system is shown after about 30000
generations in figure 4, and run to completion, this run allowed 759 audience
members to view the performance (over double the original capacity), each on
average obscured by about one other audience member. Note how upper tiers have formed above raked seating zones. In addition, the system seems to evolve
‘staircases’. Locations without a view
are unoccupied, and looking at the figure from above, it is possible to see
blocks winding up towards the higher reaches of the theatre, which once would
have been occupied, themselves, but now merely serve for access.
In the last paragraph, it was noted that we only
allowed the accessible locations to grow.
What would happen if any location were allowed to grow regardless of
location? As the system must improve
fitness through any growth, we do not see uncontrolled increase in blocks, but
in fact observe the situation shown in figure 1. Although this looks very similar to figure 4, careful inspection
of figure 1 shows that the lower tiers are flattened, without the raking
observed in figure 4. This is due to
the fact that outward growth can occur at any time, not just when a cell is at
the top level, and thus will often trigger to add another audience member, at
the cost of restricting the view of those already present.


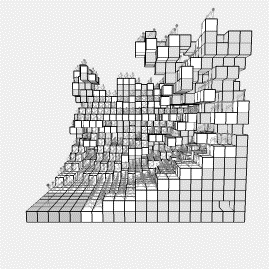

Figure
4: Four views of
the cellular growth model after about 30000 generations
One problem of the cell growth method above is that
it is an additive process. Although it
could be adapted to also remove blocks, it is difficult to do so and maintain
structural integrity. Therefore, we
turned to a few other well known techniques to attempt sparser or more regular
growth. Firstly, we implemented a
diffusion-limited aggregation method.
Cells were allowed drop from above, or fly in along one of the cardinal
axes, sticking if they collided with another block. The results were less good than the cellular block growth,
although, by ensuring the same rule of only allowing a change if the added
block improved the fitness, the actual mechanics of the two algorithms are
fairly similar: the aggregation differs in that places in the middle tend to
blocked by towers to the sides collecting material, but otherwise the growth
method tends to be based on cells sticking to other accessible (and therefore
exposed) locations.
The final two experiments were conducted using
evolutionary algorithms. These followed
standard genetic algorithm operators for mutation, crossover and selection
[14], although we also implemented a mutation only GA, essentially equivalent
to simulated annealing. The genotype
for the first evolutionary algorithm contained a string of 8000 bits – one for
every location in the 20x20x20 grid.
The bit represented block or no block, but only if the block was
reachable through others from the ground, ensuring some structure to the form. Crossover, where used, was uniform. Figure 5 shows the result of evolution using
the mutation only method, after approximately 40000 generations. As can be seen, although structure is
enforced, it can be in a snake-like pattern.
The results were not good generally, with most experiments producing
around 480 seats with an average of three intervening people. This appeared to mainly be due to the
non-structural parts, previously unseen in the aether being ‘linked’ in through
mutation, suddenly accruing a connected chain, but having no real optimisation
of the floor plain into raked seating.


Figure
5: Standard genetic
algorithm evolution
In order to overcome the problem of unstructured
seating, we applied a genetic program, evolved using similar operators to the
genetic algorithm. In this case, each
genotype consisted of 16 genes. Each
gene coded a pointer to another gene to grow in each dimension (up, down, left,
right, front, back), and a timeout, after which the gene would deactivate, in
terms of the recursion depth from the seed.
The seed was placed at 0,0,0 (the furthest corner from the stage). It quickly became apparent that the order of
applying identical genes drastically affected the outcome, and ultimately, a
fixed order of application was adopted (i.e., left then right then up, etc),
despite the fact that this would tend to skew systems to right-handedness or
left-handedness. The results near the
beginning of a run are shown in figure 6(a), and at the end in figure 6(b). Despite the regularity of the solutions and seemingly
many seats, the performance of the system was at best around 500 places, with
each audience member on average obscured by approximately 2 others.
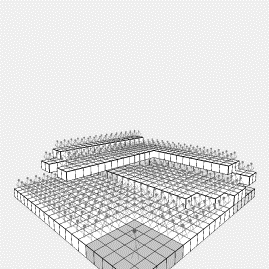
Figure
6: (a) Early run
results from the genetic program experiment (b) the same run after about 20000
generations
The genetic program suffered considerably from early
convergence, perhaps because minor gene changes make such drastic differences
to the final structure. Despite the
often tantalising range of structures shown early in the runs [see figure 6(a)]
the outcome was usually very similar: a pyramidal type object towards the rear
and no other structure. Figure 6(b)
shows about the most interesting result from the run. The genetic program often seemed stunted by its own regularity:
the requirement for accessibility necessitates single steps upwards, that often
need offsets from one row to the next.
Features such as the ‘staircases’ observed in the cellular growth models
did not occur.
4. Conclusion
In this paper we examined several algorithms to
generate an ecomorphic environment around the notion of ‘theatre’. An ecomorphic environment is one that is
structurally coupled to the activities of its occupants. In this case, the activity
is a play, where both actors and audience are engaged in an ongoing process
about their various roles in the performance, be it participant or observer, or
a combination of the two. We started
with an individualistic approach to growth of form which failed due to the
avarice of the individuals within the audience. Each individual optimised their own visibility, whilst blocking
the accessibility for other agents. In
a sense this is unsurprising, as the building growth is not about the activity
in general, but in relation to individuals within it. Thus we modified the rules to create a cellular growth model,
evaluated according to the accessibility to the population and visibility of
the stage at every accessible location. The evaluation ultimately should also
contain structural analysis of the results, although by its very nature the
cellular growth algorithm ensures constructability. The algorithm, while limited to additive growth without pruning
or rejuvenation, performed better for the numbers of audience members satisfied
than more sophisticated diffusion-limited aggregation and genetic algorithm
approaches. Its aesthetic seemingly
followed the ecomorphic intention as a randomised yet consistent growth of a
multi-tiered theatre with raked seating.
The algorithm embodies a natural evolution around the activity within
the environment from step to step. As
such it can of course only hillclimb towards a goal. If there are local maxima then the algorithm may halt
suboptimally. However, this is not as
restrictive as it may sound. It conforms
with precepts of evolutionary growth: that an intermediate step must be a
viable individual, and it does not necessarily imply a fixed output form. An analogy can be made with scaffolding,
where the structure is advantageous to get a certain amount of the way, but may
then be removed later to reveal a more appropriate form. This seems in fact to occur, with previous
seating turning to ‘staircases’ out of view of the stage when further seating
is added. Indeed, we might ask could
local optima even exist for this problem?
The complex nature of the evaluation function allows for many different
approaches to the building of form whilst still evolving a better outcome. The interdependent constraints allow an
offset of costs, so that a move that lowers fitness in one area may raise it in
another. As an exploration, therefore,
the system offers a creative process which adapts to its occupation through a
natural progression of design evolution.
5. References
1.
Turner, A., 2002: “Ecomorphic dialogues.” In C. Soddu (ed) Proceedings
Generative Art 2002 pp. 38.1–38.8
2.
Maturana, H.R. and Varela, F.J., 1979: Autopoiesis and Cognition: The
Realization of the Living,
Kluwer Academic Publishers
3.
Varela, F.J., Thompson, E. and
Rosch, E. 1993: The Embodied Mind:
Cognitive Science and Human Experience, MIT Press
4.
Thompson, D’A. W., 1961: On Growth and Form (Abridged Edition),
Cambridge University Press
5.
Grassé, P.-P. 1959: La Reconstruction
du nid et les coordinations interindividuelles. La théorie de la stigmergie,
Insectes Sociaux 6: 41-84.
6.
Luhmann, N. 1996: Social Systems, Stanford University
Press
7.
Hillier, B.
and Hanson, J. 1984: The Social Logic of
Space, Cambridge University Press, Cambridge.
8.
Soddu, C.
2004: “Generative Design / Visionary Variations - Morphogenetic processes for
Complex Future Identities” in P. Van Looke & Y. Joye Aesthetics and Generative Methods in Architectural Design, Communication & Cognition 36
9.
Coates,
P.S., Broughton, T. and Jackson, H. 1999: “Exploring Three-dimensional Design
Worlds using Lindenmayer systems and Genetic Programming” in P. Bentley (ed) Evolutionary Design by Computers
10.
Coates, P.,
Healy, N., Lamb, C. and Voon, W.L. 1996: “The use of cellular automata to
explore bottom-up architectonic rules.” Eurographics UK Chapter 14th Annual
Conference, Imperial College, London.
11.
Witten, T.A.
and Sander, L.M. 1981: Diffusion-limited aggregation, a kinetic critical
phenomenon.” Physical Review Letters 47 1400-1403.
12.
Holland,
J.H. 1975: Adaptation in Natural and Artificial Systems: An Introductory Analysis
with Applications to Biology, Control and Artificial Intelligence, University of Michigan Press
13.
Koza, J.R.
1993: Genetic Programming: On the Programming of Computers by Means of Natural
Selection, MIT Press
14.
Goldberg, D.E. 1989: Genetic
Algorithms in Search, Optimization and Machine Learning, Addison-Wesley